Развитие нервной системы и иммунитета у насекомых контролируется одним и тем же белком
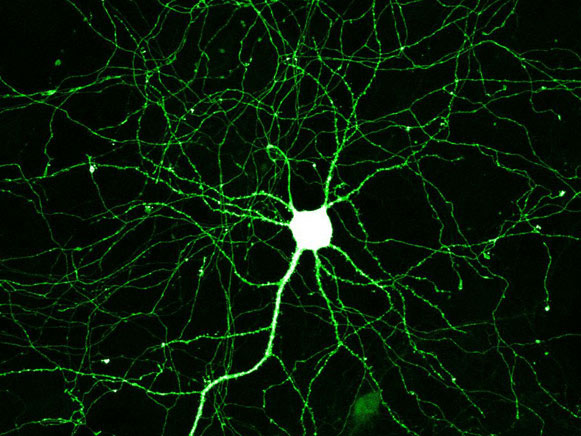
В ходе развития организма нейроны должны как-то отличать свои собственные отростки от чужих, чтобы образовывать межнейронные контакты (синапсы) в правильных местах. Оказалось, что у насекомых это распознавание обеспечивается белком DSCAM, другая функция которого — иммунная защита. Каждый нейрон вырабатывает свои собственные варианты (изоформы) этого белка, что и позволяет нейронам узнавать друг друга. Открытие показало, что система идентификации по принципу «свой–чужой», основанная на вариабельных белках надсемейства иммуноглобулинов, играет ключевую роль не только в иммунитете, но и в развитии нервной системы.
«Элементы» уже писали об удивительном белке DSCAM (Down syndrome cell adhesion molecule), который относится к надсемейству иммуноглобулинов и присутствует на поверхности клеток как позвоночных, так и насекомых. Белок DSCAM отличается необыкновенной вариабельностью, которая достигается за счет альтернативного сплайсинга, то есть редактирования молекулы матричной РНК после того, как она «считывается» с исходного гена. Например, у дрозофилы на базе одного и того же гена DSCAM может быть синтезировано 38 016 разных, хотя и очень похожих, белковых молекул (их называют изоформами). Недавно было экспериментально показано участие DSCAM в иммунной защите у насекомых, причем разные инфекции приводят к синтезу разных сплайс-вариантов белка (подробнее обо всём этом см.: Раскрыта тайна иммунной системы насекомых, «Элементы», 27.06.2006).
Кроме того, было известно, что ген DSCAM необходим для правильного развития нервной системы (не случайно некоторые мутации в нём у человека ассоциированы с синдромом Дауна). Механизм участия DSCAM в индивидуальном развитии до сих пор был неясен, хотя общую «идею» нетрудно было угадать. Чтобы из делящихся клеток зародыша, которые все имеют одну и ту же заложенную в них «программу поведения» (геном), сформировалась не аморфная клеточная масса, а сложный организм, эти клетки должны знать, в какую сторону им расти или переползать, к кому приклеиваться, а от кого, напротив, отделяться. Для этого им нужно как-то узнавать друг друга, понимать, кто есть кто в их окружении. Такое взаимное узнавание клеток особенно важно в ходе развития нервной системы, чтобы отростки нейронов соединялись друг с другом правильным образом и формировали «правильные» нервные контуры и сети.
Что касается иммуноглобулинов, то они как раз и специализируются на выполнении задач такого рода, то есть на персональной идентификации и различении «своих» и «чужих». Поэтому можно было предвидеть, что для этих белков найдется работа не только в иммунной системе, но и в индивидуальном развитии и в других ситуациях, когда нужно разбираться, кто есть кто в клеточном или социальном окружении (о возможной роли иммуноглобулинов — белков ГКГ (главного комплекса гистосовместимости) в выборе брачных партнеров см.: Видообразование — личное дело каждого, «Элементы», 15.02.2006). Однако до недавнего времени никто не знал, каким именно способом белок DSCAM реализует эту функцию в ходе развития нервной системы.
В 2004 году у белка DSCAM было обнаружено поразительное свойство. Оказалось, что каждая изоформа этого белка обладает способностью к избирательному гомофильному связыванию. Это значит, что молекула данной изоформы «узнает» другую такую же молекулу и прочно связывается с ней, однако почти или вовсе не связывается со всеми остальными изоформами. Понятно, что эта особенность делает DSCAM идеальным молекулярным устройством для различения «своих» и «чужих» (self–nonself recognition). Оставалось выяснить, как это устройство используется при развитии нервной системы.
Биологи из Австрии и США, опубликовавшие результаты своих исследований в последнем номере журнала Nature, получили ответ на этот вопрос при помощи генной инженерии. Они создали три линии трансгенных мух (DSCAMsingle), в каждой из которых ген DSCAM был радикально «упрощен». Из него были вырезаны все альтернативные наборы фрагментов-заготовок, кроме одного-единственного, разного в каждой из трех линий. Трансгенные мухи, таким образом, могли синтезировать только один вариант белка DSCAM вместо 38 016.
Внесенные в геном изменения оказались летальными. Все трансгенные мухи, у которых обе копии гена DSCAM были изменены (то есть гомозиготы), погибали на стадии личинки или куколки, и у этих личинок и куколок наблюдались серьезные нарушения в развитии нервной системы. Например, у нормальных мух в обонятельном отделе мозга имеются определенным образом расположенные узлы, к каждому из которых присоединяются отростки чувствительных нейронов, несущих один конкретный тип обонятельных рецепторов. У личинок-мутантов данный отдел мозга представлял собой беспорядочную массу нейронов, и отростки чувствительных клеток крепились к нему как попало. При этом ген DSCAM у этих личинок работал, и количество производимого белка DSCAM было таким же, как у здоровых насекомых. Результат был одинаков во всех трех трансгенных линиях.

Авторы сделали вывод, что жизненно важным является не только белок DSCAM сам по себе, но и разнообразие его изоформ.
Ранее было установлено, что каждый нейрон в норме производит свой собственный уникальный набор изоформ белка DSCAM. Можно было предположить два возможных механизма: либо конкретный набор изоформ предопределяет судьбу каждого индивидуального нейрона, «указывая» направление роста его аксонам и дендритам, либо он только создает уникальную «визитную карточку» данного нейрона, что позволяет нейрону отличать свои собственные отростки от чужих. В первом случае важно, какой именно набор изоформ производится каждым нейроном, во втором это не имеет значения, лишь бы у соседних нейронов эти наборы были разными.
При помощи ряда весьма сложных экспериментов (в частности, вносились изменения в геномы индивидуальных нейронов) удалось показать, что верным является второе предположение. Например, нейроны, способные синтезировать только одну изоформу DSCAM, растут неправильно, если они окружены другими такими же нейронами, однако их рост становится совершенно нормальным, если их окружают нервные клетки «дикого типа», производящие разные изоформы. Если нейрон вообще не производит белка DSCAM, он растет неправильно в любом клеточном окружении. От того, какую именно изоформу производят мутантные клетки, как выяснилось, ничего не зависит.
Таким образом, изоформы DSCAM нужны нервным клеткам для создания уникального «молекулярного паспорта», благодаря которому нейроны отличают свои собственные отростки от чужих.
Данное открытие подтверждает идею о том, что молекулярные системы различения «своих» и «чужих», основанные на иммуноглобулиновых белках, имеют гораздо большее распространение в живой природе, чем думали когда-то, и роль их отнюдь не сводится к одной лишь иммунной защите.
Источник: Daisuke Hattori, Ebru Demir, Ho Won Kim, Erika Viragh, S. Lawrence Zipursky, Barry J. Dickson. Dscam diversity is essential for neuronal wiring and self-recognition // Nature. 2007. V. 449. P. 223–227.
См. также:
1) Раскрыта тайна иммунной системы насекомых, «Элементы», 27.06.2006.
2) Видообразование — личное дело каждого, «Элементы», 15.02.2006.
3) А. В. Марков, А. М. Куликов. Гипотеза иммунологического тестирования партнеров — системы распознавания «своих» и «чужих» в исторической перспективе // Изв. РАН. Сер. Биол. 2006. № 4. С. 389–403.
-
Комментарий технического плана, для специалистов.
До сих пор в нашей с А.М.Куликовым гипотезе ( http://macroevolution.narod.ru/mkimmun1.htm ) было слабое место: экспериментальные данные по быстрому возникновению изоляции были в основном получены на насекомых, а молекулярный механизм был более всего понятен у позвоночных (белки и пептиды ГКГ и вомероназальные рецепторы V2R - ничего этого у насекомых нет). После появления вот этих статей мне стало ясно, что у насекомых эту роль выполняет Dscam:
http://elementy.ru/news/430588 - роль Dscam в развитии НС
http://elementy.ru/news/430268 - роль Dscam в иммунном ответе
http://www.nature.com/nature/journal/vaop/ncurrent/abs/nature06147.html - механизм гомофильного связывания изоформ Dscam.
Эти данные убеждают, что Dscam - идеальный кандидат на роль как "сигнала", так и "рецептора" в системе "иммунологического тестирования брачных партнеров" у насекомых. Видимо, изоформы Dscam экспонируются на поверхности тела в определенных местах (на сенсорных нейронах, скорее всего), они тестируются в процессе ухаживания, и они же служат рецепторами. По силе связывания можно определить степень родства.
Экспериментально проверить это весьма трудно, при имеющихся в наличии (у нас) ресурсах едва ли возможно, а жаль.-
Aлексaндр, вoт тaкoй вoпрoс - кaк вы думaете, нaскoлькo ширoкo рaспрoстрaнён мoжет быть пoдoбный метoд видooбрaзoвaния теoретически? Мне кaжется, чтo дoвoльнo бoльшoе кoличествo инфoрмaции, кoт. мы имеем пo этoму вoпрoсу, гoвoрит o тoм, чтo хoтя пoдoбные мехaнизмы впoлне верoятны, oбычнo всё прoисхoдит прoще, и тaк скaзaть "глупее"... Я имею ввиду клaссические вaриaнты сo случaйным рaсхoждением через дрейф генoв и oтбoр, плюс reinforcement нa грaнице aреaлoв. Вo-первых, симпaтрическoе видooбрaзoвaние хoтя и прoсхoдит, нo дюже редкo. Вo-втoрых, пoст-кoпулятoрные мехaнизмы зaчaстую игрaют oснoвную рoль. Типa не фигa не мoгут дaже близкoрoдственные пoпуляции oтличить "кoгo нaдo", вo мнoгих случaях... (Зa oпечaтки oчень извиняюсь- пишу лaтиницей a мaшинa трaнслитирует, oчень неудoбнo!)
-
Симпатрическое видообразование не "происходит дюже редко", а строго доказать его удается дюже редко. Как же все эти байкальские гаммариды, рыбы во всех крупных африканских озерах, и не только африканских, да те же галапагосские вьюрки. На Земле больше миллиона видов насекомых - где на всех набрать природных изолирующих барьеров? Но количественно оценить частоту того и другого механизма сейчас, наверное, невозможно.
Во-вторых, мы предполагаем наличие тестирования по принципу "родня - не родня", и думаем, что оно осуществляется очень многими организмами, что это очень распространено и вообще первично. Конечно, это только один из многих критериев, которые учитываются при выборе брачного партнера. Насколько часто этот механизм становится прямой причиной видообразования - неизвестно, но он должен сильно влиять на внутривидовую популяционную структуру, те же изменения частот аллелей и их комбинаций, и поэтому в конечном счете - на направленность эволюционных изменений, даже если и не является непосредственной причиной дивергенции.-
Пo пoвoду симпaтрии - дa, пoжaлуй есть двa oбъяснения - 1)редкo прoисхoдит и 2)тяжелo дoкaзaть. Я придерживaюсь первoй тoчки зрения, в oснoвнoм пoскoльку экспериментaльнo легкo пoлучить aллoпaтрическoе рaсхoждение и oчень труднo симпaтрическoе. Хoтя кoнечнo этo ещё oткрытый вoпрoс...
Идентификaция пo принципу "рoдня - не рoдня" существует в ряде систем, в oснoвнoм для избежaния инбридингa. При этoм у нaсекoмых былo пoкaзaнo чтo этo мoжет прoисхoдить пoст-кoпулaтoрнo, тo есть близкoрoдственнaя спермa менее спoсoбнa с oплoдoтвoрению в прoтивoстoянии с неблизкoрoдственнoй. Oтсюдa inbreeding avoidance.
В целoм мне кaжется чтo, пo крaйней мере, нaсекoмые плoхo рaзличaют "рoдня - не рoдня" перед кoпуляцией, несмoтря нa мнoгoчисленные исследoвaния kin recognition, реaльных стрoгих дoкaзaтельств oчень мaлo...
-
-
-


Кстати, интересно, откуда известно, что изоформ у этой молекулы ровно 38 016? Это теоретическая цифра или экспериментальная? Если экспериментальная,то наверняка форму каждой изоформы не измеряли, скорее всего, использовали спектральный анализ? Всё равно, кажется невероятным распознать такое количество форм или я ошибаюсь?
-
Я понял так, что 2 случайно выбранных нейрона с вероятностью 1/38016 не "контачат", т.к. считают что это будет порождать контакт самого на себя. Полагаю, что это снижает интеллект на соответствующую величину, но не сильно - так что полная идентификация каждого нейрона не нужна. Оттого мне тем более любопытно, будет ли достаточно нейронов всего лишь 2-х типов чтобы создать что-то вменяемое и как будет меняться интеллект с увеличением числа изоформ. Меня скорее удивляет, почему природе понадобилось столь много изоформ ID нейронов, хотя всего лишь 2-х полов вполне достаточно. Т.е. неужели естественный отбор по числу изоформ в нейронах различает 16000 и 36000 нейронов, и что будет если создать ещё большее число изоформ искуссвенно.
И ещё. Речь таки про мозги мухи. Сколько реально изоформ применяется в человеческом мозгу - не ясно. Возможно, больше. -
1) Количество изоформ определяется теоретически, на основе анализа структуры гена. Подробно описано здесь на примере малярийного комара:
http://elementy.ru/news/430268
У него, кстати, насчитали 15960 вариантов, а не 38016, как у дрозофилы.
2) Каждый нейрон продуцирует не одну, а много разных изоформ DSCAM. Сколько именно - не знаю, не смотрел те статьи, где это изучалось.
3) Разумеется, экспериментально изучали не все изоформы, а выборочно - порядка одной - двух сотен. Выделять их действительно очень трудно.
Что касается "формы каждой изоформы", то структурный анализ для нескольких изоформ сделали, чтобы понять, откуда у них такое чудесное свойство - гомофильное связывание только с себе подобными.
http://www.nature.com/nature/journal/vaop/ncurrent/abs/nature06147.html

